From the Access Research Network at www.arn.org/docs/mm/cilium_all.htm
Molecular Machines Museum
The Cilium
|

|
The cilium is
an example of what Michael Behe describes as an irreducibly complex
system. In his book, Darwin's Black Box, he explains that such
irreducibly complex systems could not have arisen by a gradual
step-by-step Darwinian process.
|
|

|
In many
biological structures proteins are simply components of larger molecular
machines. Like the picture tube, wires, metal bolts and screws that
comprise a television set, many proteins are part of structures that
only function when virtually all of the components have been assembled.
A good example of this is a cilium. Cilia are hairlike organelles on the
surfaces of many animal and lower plant cells that serve to move fluid
over the cell's surface or to "row" single cells through a
fluid. In humans, for example, epithelial cells lining the respiratory
tract each have about 200 cilia that beat in synchrony to sweep mucus
towards the throat for elimination.
|
|
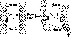
|
A cilium
consists of a membrane-coated bundle of fibers called an axoneme. An
axoneme contains a ring of 9 double microtubules surrounding two central
single microtubules. Each outer doublet consists of a ring of 13
filaments (subfiber A) fused to an assembly of 10 filaments (subfiber
B). The filaments of the microtubules are composed of two proteins
called alpha and beta tubulin. The 11 microtubules forming an axoneme
are held together by three types of connectors: subfibers A are joined
to the central microtubules by radial spokes; adjacent outer doublets
are joined by linkers that consist of a highly elastic protein called
nexin; and the central microtubules are joined by a connecting bridge.
Finally, every subfiber A bears two arms, an inner arm and an outer arm,
both containing the protein dynein.
|
|



|
But how does a
cilium work? Experiments have indicated that ciliary motion results from
the chemically-powered "walking" of the dynein arms on one
microtubule up the neighboring subfiber B of a second microtubule so
that the two microtubules slide past each other. However, the protein
cross-links between microtubules in an intact cilium prevent neighboring
microtubules from sliding past each other by more than a short distance.
These cross-links, therefore, convert the dynein-induced sliding motion
to a bending motion of the entire axoneme.
Now, let us sit
back, review the workings of the cilium, and consider what it implies.
Cilia are composed of at least a half dozen proteins: alpha-tubulin,
beta-tubulin, dynein, nexin, spoke protein, and a central bridge
protein. These combine to perform one task, ciliary motion, and all of
these proteins must be present for the cilium to function. If the
tubulins are absent, then there are no filaments to slide; if the dynein
is missing, then the cilium remains rigid and motionless; if nexin or
the other connecting proteins are missing, then the axoneme falls apart
when the filaments slide.
|
|

|
What we see in
the cilium, then, is not just profound complexity, but also irreducible
complexity on the molecular scale. Recall that by "irreducible
complexity" we mean an apparatus that requires several distinct
components for the whole to work. A mousetrap must have a base, hammer,
spring, catch, and holding bar, all working together, in order to
function. Similarly, the cilium, as it is constituted, must have the
sliding filaments, connecting proteins, and motor proteins for function
to occur. In the absence of any one of those components, the apparatus
is useless.
|
|
|
The components
of cilia are single molecules. This means that there are no more black
boxes to invoke; the complexity of the cilium is final, fundamental. And
just as scientists, when they began to learn the complexities of the
cell, realized how silly it was to think that life arose spontaneously
in a single step or a few steps from ocean mud, so too we now realize
that the complex cilium can not be reached in a single step or a few
steps. But since the complexity of the cilium is irreducible, then it
can not have functional precursors. Since the irreducibly complex cilium
can not have functional precursors it can not be produced by natural
selection, which requires a continuum of function to work. Natural
selection is powerless when there is no function to select. We can go
further and say that, if the cilium can not be produced by natural
selection, then the cilium was designed.
Other examples
of irreducible complexity abound, including aspects of protein
transport, blood clotting, closed circular DNA, electron transport, the
bacterial flagellum, telomeres, hotosynthesis, transcription regulation,
and much more. Examples of irreducible complexity can be found on
virtually every page of a biochemistry textbook.
|
|
|
From
"Molecular Machines: Experimental Support for the Design
Inference" by Michael J. Behe.
|
Copyright © 1998 Michael J. Behe. All rights reserved.
International copyright secured.
File Date: 6.10.98
This data file may be reproduced in its entirety
for non-commercial use.
A return link to the Access Research Network web site would be appreciated.
Page development by Premier Publications
for Access Research Network,
Colorado Springs, Colorado, USA.
|
